|
Weevil News
|
http://www.curci.de/Inhalt.html
|
No. 37
|
10 pp.
|
11th September 2007
|
ISSN 1615-3472
|
|
Bayer, C., H. Winkelmann & F. Bahr (2007):
Ergebnisse einer faunistischen Studie auf der Insel Rhodos. Erster
Beitrag zur Fauna von Griechenland (Coleoptera, Curculionoidea). -
Weevil News: http://www.curci.de/Inhalt.html, No. 37: 10 pp., CURCULIO-Institute: Mönchengladbach. (ISSN 1615-3472).
|
Ergebnisse einer faunistischen Studie auf der Insel Rhodos.
Erster Beitrag zur Fauna von Griechenland
(Coleoptera: Curculionoidea).
von
Christoph Bayer (Berlin), Herbert Winkelmann
(Berlin) und Friedhelm Bahr (Viersen)
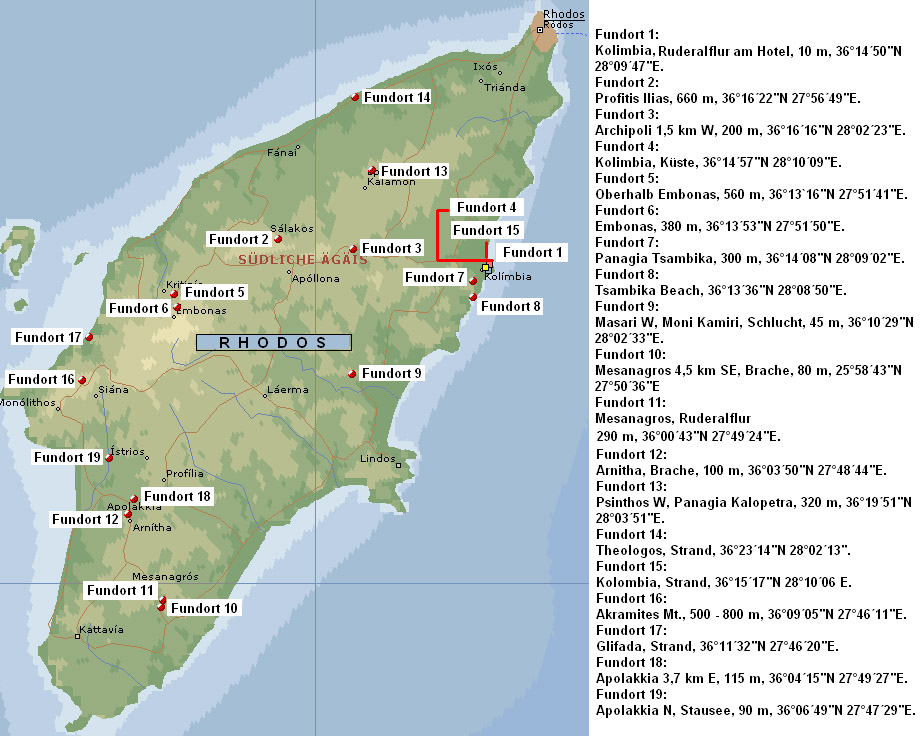
Mit 152 Abbildungen auf 37 Bildtafeln, 1 Karte und 1 Tabelle.
Manuscript received: 28th July 2007
Accepted: 2nd September 2007
Abstract:
Results of a faunistic study on the island of Rhodes. First
contribution to the fauna of Greece (Coleoptera: Curculionoidea).
This contribution to the weevil fauna of Rhodes (Greece) contains a
description of 19 localities, species lists and numerous pictures of
biotopes and specimens. The data were collected during a field trip
from 31st March to 14th April 2007 to Rhodes. The
species list of Rhodes will be part of the online-catalogue "Weevil
Fauna of Greece" which will appear in 2007. It will refer also to
problems of determination and taxonomy of the weevils of this region.
Zusammenfassung
Im Rahmen der Erforschung der Rüsselkäfer-Fauna
Griechenlands wurde vom 31.3. bis 14.4.2007 eine Exkursion nach Rhodos
unternommen. Es wurden 19 Standorte untersucht. Dieser Beitrag
enthält eine Karte der Fundorte, zahlreiche Fotografien der
Biotope, Pflanzen- und Lebendaufnahmen ausgewählter
Rüsselkäfer-Arten. Die Gesamtartenliste der bisher
nachgewiesenen Arten von Rhodos wird in den in Kürze erscheinenden
Katalog der Rüsselkäfer Griechenlands übernommen.
1. Einleitung
Im Rahmen der Untersuchungen der
Rüsselkäferfauna Griechenlands unternahmen die Autoren vom
31.3. - 14.4. 2007 eine Exkursion zur Insel Rhodos. Die Insel liegt in
der Ost-Ägäis und ist die Hauptinsel der Inselgruppe Dodekanes
("Zwölf Inseln"). Rhodos ist ca. 80 km lang und 35 km breit, die
Gesamtfläche beträgt fast 1400 km². Sie ist die
südöstlichste Insel Griechenlands und liegt nur knapp 20 km
südlich des türkischen Festlands [W37.1].
Flora und Fauna weisen daher Beziehungen zu Kleinasien auf. Wir
rechneten daher damit, auf viele Arten zu stoßen, deren
Hauptvorkommen auf dem türkischen Festland liegt. Auf Rhodos
liegen mehrere kleine Bergketten, deren höchste der 1215 m hohe
Attaviros ist. Die Gipfel der weiteren Berge liegen bei 600-800 m
Höhe. Aufgrund der geringen Höhe der Berge und der Nähe
zum Festland ist – bezogen auf die Rüsselkäferfauna
– das Vorkommen von Endemiten auf Rhodos eher unwahrscheinlich.
Da wir für unsere erste Erkundung der Insel das zeitige
Frühjahr gewählt haben, konnten wir noch die
Hauptblütezeit der mediterranen Thero- und Geophyten erleben [W37.2].
Besonders die Formenvielfalt und Farbenpracht der Orchideen war
beeindruckend, auch wenn diese Pflanzenfamilie im mediterranen Raum von
Phytophagen kaum genutzt wird [W37.3].
Floristisch weist Rhodos hingegen einige Besonderheiten auf. Neben dem
Vorkommen von Endemiten ist auch das Fehlen bestimmter Taxa, das erst
in neuerer Zeit systematisch untersucht wird, ein interessantes und oft
noch nicht erklärtes Phänomen. So fehlt auf Rhodos
beispielsweise die Gattung Viola (Veilchen). Auf dem nahe gelegenen türkischen Festland jedoch kommen Viola-Arten
wie auch im übrigen Mediterrangebiet und auf vielen Inseln der
Ägäis vor. Bei der gezielten Suche nach annuellen
Entwicklungspflanzen in Therophytenfluren und auf Ackerbrachen fiel uns
darüber hinaus das Fehlen einjähriger Veronica-Arten (Ehrenpreis) auf. Veronica anagallis-aquatica L. (Blauer Wasser-Ehrenpreis) war die einzige Art der Gattung auf dieser Insel [W37.4]. Ebenfalls bemerkenswert ist die Arten- und Individuenarmut der Gattung Verbascum
(Königskerze), da das türkische Festland ein Schwerpunkt der
Formenvielfalt dieser Gattung ist. Es verwundert daher nicht, dass kein
Vertreter der Gattungen Cionus, Cleopus und Gymnetron (Rhinusa, pars) gefunden wurde, was jedoch auch jahreszeitlich bedingt sein könnte, da die Arten der Verbascum-Zönose
erst zu Beginn der Blütezeit der Königskerze ohne
größeren Aufwand nachgewiesen werden können.
Schließlich wurden bei der gezielten Suche nach
Rüsselkäfern auch andere interessante Tiere beobachtet [W37.5].
Da die Autoren erstmalig Rhodos besuchten
und nur zwei Wochen Zeit hatten, sollten möglichst
unterschiedliche Standorte von der Küste bis zu den höchsten
Bergen untersucht werden, um einen ersten Überblick über das
Spektrum der Lebensräume zu erhalten. Von einer günstigen
Unterkunft bei Kolimbia (Nordostküste) wurden sowohl Biotope in
der direkten Umgebung (Fundorte 1, 4, 7, 8, und 15) [W37.6] als auch weiter entfernte Fundorte aufgesucht. Typische Küstenstandorte waren die Fundorte 4, 8, 14, 15 und 17 [W37.7], typische Bergstandorte die Fundorte 2 und 16 [W37.8].
Das einzige größere Süßgewässer auf Rhodos
ist ein Stausee im Süden der Insel, dessen Uferbereich bei einem
natürlichen Zulauf untersucht wurde (Fundort 19) [W37.9]. Bei Fundort 11 handelt es sich um eine typische Ruderalflur am Rand einer Siedlung [W37.10]. Fundort 9 ist eine breite Schlucht mit einem flachen, trocken gefallenen Flussbett [W37.11].
3. Bemerkenswerte Beobachtungen
Das Besondere an einer faunistischen
Exkursion ist für uns die Möglichkeit, Beobachtungen zur
Lebensweise und zum Verhalten der Tiere zu machen. Diese werden im
besten Fall als Fotografie oder Videosequenz festgehalten. Wer den
Aufwand kennt, der für den Nachweis der meisten
Rüsselkäfer-Arten erforderlich ist, kann nachvollziehen, dass
allein die Bilddokumentation der Beobachtungen im Gelände eine
gewisse Herausforderung darstellt. Dies trifft umso mehr zu, wenn
bestimmte Qualitätsstandards eingehalten werden sollen. Lebendabbildungen
von Rüsselkäfern und ihren Entwicklungsstadien sind für
uns von großer Bedeutung, sagen sie doch zur Lebensweise der Art
sehr viel mehr aus als die klassische Dorsalabbildung eines oft
unnatürlich präparierten Sammlungsexemplars. So zeigen
Lebendabbildungen oft erst die arttypische Färbung einer Imago und
damit deren Tarnung im Biotop oder weitere spezielle Anpassungen.
Beispiele hierfür sind die Wachsüberzüge vieler
Cleoninae oder auffällige Schuppenmuster sowie der "Mantel aus
Erdreich" vieler bodenlaufaktiver Arten, deren Tarnung und
Totstellverhalten (Thanatose) mit Hilfe von Lebendfotos aus dem
natürlichen Lebensraum gut nachvollziehbar wird [W37.12]. Darüber hinaus verdeutlichen auch Abbildungen von Aufenthalts-, Fraß- und Entwicklungspflanzen
viele Aspekte der Überlebensstrategien phytophager Käfer. Sie
zeigen z. B. den Entwicklungszustand der Pflanze bei der Eiablage und
während der Larvalentwicklung oder machen dem Betrachter die Wahl
des Verpuppungsortes unter dem Aspekt des Schutzes und der Tarnung
verständlich [W37.13]. Da es am Fundort zeitlich nicht möglich ist, die gesamte Entwicklung und Biologie einer Art zu beobachten, liefern Fütterungs- und Aufzuchtversuche
wertvolle Informationen. Auch die im Gelände nicht leicht zu
beobachtende Verpuppungs- und Schlupfphase der Tiere kann dann im
Detail untersucht und dokumentiert werden [W37.14].
Schließlich sei noch auf die
überwiegende Zahl der Arten hingewiesen, zu denen keine
"bemerkenswerten Beobachtungen" gemacht werden konnten. Die Arten
dieser "Kategorie" wurden entweder mittels "unspezifischer" Methoden
(Keschern, Klopfen, Sieben …) oder bei der gezielten Suche oder
der Beobachtung anderer Arten "nebenbei" entdeckt, und landen somit auf
der Artenliste [W37.15].
Oft sind dies migrierende Einzelexemplare oder bodenlaufaktive Arten
(von denen viele zu den so genannten "unspezialisierten
Kurzrüsslern" gehören, deren Larven sich frei im Boden an den
Wurzeln ihrer Wirtspflanzen entwickeln). Bei diesen Arten Eiablage oder
Larvalentwicklung zu studieren, gehört zu den größeren
Herausforderungen and den Biologen und nimmt viel Zeit in Anspruch.
Daher ist diese Artengruppe auch etwas Besonderes, denn die
Klärung ihres Verhaltens und ihrer Ansprüche an den
Lebensraum gehören auf unsere Agenda!
Zur Verdeutlichung des unterschiedlichen
Wissensstandes, der zu den einzelnen Arten existiert, sollen hier drei
"Erforschungsgrade" verglichen und mit Beispielen von der Insel Rhodos
belegt werden:
1. Gut untersuchte Arten: Lebensweise, Entwicklungspflanzen und Lebenszyklus
sind hinreichend bekannt (meist so genannte "Schädlinge" an Nutzpflanzen)
[W37.16].
2. Unzureichend gut untersuchte Arten: Entwicklungspflanze oder einige Fraß-,
beziehungsweise Aufenthaltspflanzen bekannt. Daten zur Entwicklung und zu
den Ansprüchen an den Lebensraum fehlen oder sind nur lückenhaft belegt
[W37.17].
3. Nicht untersuchte Arten: Daten zu Lebensweise, Entwicklungspflanzen und
Lebenszyklus fehlen weitgehend oder basieren auf Zufallsbeobachtungen
[W37.18].
Kommentare zu ausgewählten Taxa:
Bagous sp.
Die artenreiche Küstenvegetation an Fundort 15 wurde bei
mehreren Begehungen intensiv abgesucht. Durch Abklopfen der
Krautschicht wurde auch ein Männchen der Gattung Bagous gefunden. Ein Zusammenhang mit einem typischen Bagous-Biotop
oder einer Entwicklungspflanze konnte nicht hergestellt werden. Sowohl
an Fundort 15 (schlammige Tümpel im Mündungsbereich eines
Flusses) als auch an Fundort 19 (flache Uferzone eines Stausees) wurde
gezielt nach Arten der Gattung gesucht. Diese Versuche blieben jedoch
erfolglos [W37.19].
Brachycerus spp.
In verschiedenen felsigen Biotopen wurden am Boden bereits gestorbene und in den Boden vergraben lebende Imagines der Gattung Brachycerus
gefunden. Offenbar waren die vergrabenen Tiere bereits in die
Sommerruhe gegangen, da die Imagines im Mittelmeerraum oft winteraktiv
sind. Um zu prüfen, ob die Eiablage an Zwiebelgewächsen
bereits stattgefunden hatte, wurden einige Zwiebeln z. B. der Gattung Ornithogalum (Milchstern) ausgegraben. Die Beobachtungen hierzu sind noch nicht abgeschlossen [W37.20].
Chiloneus jonicus Kraatz 1859
Dieser bodennah lebende Kurzrüssler wurde an Fundort 2 und 16
zwischen 600 und 800 m Höhe gefunden. Es handelte sich
ausschließlich um überwinterte Alt-Imagines. Aufgrund der
wenigen Exemplare, die noch am Leben waren (am Boden wurden zahlreiche
Überreste verstorbener Tiere gefunden), gehen wir davon aus, dass
die neue Generation erst im Mai erscheint. Chiloneus jonicus wird oft in Verbindung mit Urginea maritima (L.) Baker (Meerzwiebel) bzw. Asphodelus-Arten (Affodill) gefunden. Eine Entwicklungspflanzenbindung ist noch nicht bekannt [W37.21].
Coniatus suavis Gyllenhal 1834
Nach Imagines der Gattung Coniatus wurde immer wieder
erfolglos gesucht, bis an einem einzigen von Hunderten untersuchter
Bäume und Sträucher der Gattung Tamarix (Tamariske) zahlreiche gegitterte Kokons gefunden wurden [W37.23]. Aus diesen schlüpften erst nach über einer Woche Imagines von Coniatus suavis.
Das Fehlen von Imagines zu diesem Zeitpunkt des Jahres hängt
möglicherweise nur zum Teil mit der Larvalphase im Frühjahr
zusammen. Wir vermuten, dass mehrere Generationen im Jahr möglich
sind und so in der Winterpause entstandene Populationseinbrüche
kompensiert werden können. Nach Coniatus tamarisci (Fabricius 1787) wurde vergeblich gefahndet.
Donus audax Faust 1887
Um weitere Erkenntnisse zu den Entwicklungspflanzen der Hyperini zu
erhalten, wurde bei der Suche am Boden und in der Krautschicht
besonders auf Fraßspuren an Pflanzen und Aufenthaltsorte der
Larven geachtet. An Fundort 2 konnten schon Anfang April
größere Hyperini-Larven festgestellt werden, die mit Trifolium stellatum-Beständen assoziiert waren. Fraßtests bestätigten Trifolium stellatum L. in diesem Biotop als die Entwicklungspflanze, von der sich die Larven und auch die frisch geschlüpften Imagines von Donus audax
ernährten. Wahrscheinlich gehören an anderen Fundorten auch
weitere Fabaceae (Schmetterlingsblütler) zum Spektrum der
Entwicklungspflanzen [W37.24].
Donus orientalis Capiomont 1867
Bei der Suche nach Donus-Arten der höheren Lagen wurden
an Fundort 2 und 16 in 600-800 m Höhe zahlreiche Larven und auch
"geschlossene" Kokons gefunden, die sich deutlich von den Gitterkokons
von Donus audax unterschieden. Die geschlüpften Imagines konnten als Donus orientalis
bestimmt werden. An den Fundorten wurden gleichzeitig mit den Larven
und Kokons auch Imagines der überwinterten und der neuen
Generation gefunden. Den Larven wurde in den Zuchtgefäßen
eine "Salatmischung" verschiedener krautiger Pflanzen als Nahrung
angeboten. Sie fraßen ausschließlich
"Löwenzahn"-Blätter und wurden daraufhin weiterhin nur mit Taraxacum
(der mitteleuropäische Löwenzahn) gefüttert. Diese
Nahrung wurde mehrere Wochen erfolgreich angeboten, bis die Imagines
ausgehärtet waren und zur Diapause (Sommerruhe) übergingen.
Mit Donus orientalis ist ein Zuchtversuch geplant [W37.25].
Hadroplontus trimaculatus (Fabricius 1775)
Die Art kommt im südlichen Mitteleuropa, Vorderasien und
Nordafrika vor. Als Entwicklungspflanzen nennt Dieckmann [Dieckmann
1972] Cirsium vulgare (Savi) Ten. (als Cirsium lanceolatum L.) und mehrere Carduus-Arten
(Disteln, Asteraceae). An Fundort 15 konnten ein frisch
geschlüpftes Exemplar und weitere Imagines der neuen Generation
aus dem Boden unter einer Kugeldistel (Echinops sp.) ausgegraben
werden. Da die Larven vieler Arten der Ceutorhynchinae zur Verpuppung
ihre Entwicklungspflanze verlassen und im Boden einen Erdkokon bauen,
ist der Fund immaturer Tiere im Boden ein plausibler Nachweis der
Entwicklungspflanze. Es ist jedoch wahrscheinlich, dass der Kreis
potentieller Wirtspflanzen von Hadroplontus trimaculatus noch größer ist [W37.26].
Hypera jucunda (Capiomont 1868)
Auf den Sandbiotopen an der Küste (Fundort 15) wurden mehrere
immature Imagines der Art gefunden. Larven, die am gleichen Ort
gesammelt wurden entwickelten sich zu Imagines von Hypera jucunda bzw. Hypera postica.
Da die Larven beider Arten nicht zu trennen waren und sie sich bereits
kurz vor der Verpuppung befanden, konnten keine Versuche zur Bindung an
spezielle Entwicklungspflanzen gemacht werden. Die Tiere entwickeln
sich jedoch mit Sicherheit an krautigen Schmetterlingsblütlern
(Fabaceae) und wurden am Fundort oft in Beständen von Lotus halophilus
Boiss. & Spruner nachgewiesen. Für die Beobachtung der Larven
an den Entwicklungspflanzen war der Beobachtungszeitraum folglich
bereits zu spät gewählt. Wenn (und dies gilt für alle
Hyperini) die Larven des letzten Stadiums ihre Fraßpflanze
– auf der Suche nach einem geschützten Ort für den Bau
des Kokons – bereits verlassen haben, bleibt dem Beobachter nur
das Warten auf die frisch geschlüpften Tiere, deren
Fraßwahlverhalten nicht unbedingt mit dem der Larve
übereinstimmt. Daher können wir hier noch keine Angaben
über das Spektrum der Wirtspflanzen von Hypera jucunda machen [W37.27].
Hypera lunata Wollaston 1854
Die Art ist im Mittelmeerraum weit verbreitet und entwickelt sich an verschiedenen Arten der Gattung Erodium (Reiherschnabel).
Während wir an Fundort 16 auf 800 m Höhe noch Larven finden
konnten, wurden in den Sandbiotopen der Küste (Fundort 15) Kokons
mit Puppen und bereits geschlüpfte Imagines der neuen Generation
nachgewiesen. Dabei erwiesen sich sowohl die Kokons als auch die Tiere
als perfekt getarnt, so dass erstere nur durch Zufall (aufgerissener
Kokon) und letztere nur nach Ablauf ihrer Totstellzeit entdeckt werden
konnten. Erst eine Aufnahme einer Imago im natürlichen Lebensraum
macht die sonst nur als "Laune der Natur" erscheinende markante
Zeichnung des Integuments als Anpassung zur Tarnung verständlich [W37.28]. Ähnlich gezeichnet ist die nächstverwandte Art Hypera dauci
(Olivier 1807), deren Verbreitungsschwerpunkt im südlichen
Mitteleuropa liegt. Um Missverständnissen vorzubeugen, sei hier
erneut darauf hingewiesen, dass die Synonymie von Hypera fasciculata (Herbst 1795) mit Hypera lunata
oft übersehen wird, weshalb beide Namen auch in "aktuellen"
Faunenlisten oft noch nebeneinander aufgeführt werden [Alziar
2007] [Abbazzi 1994].
Hypolixus ornatus (Reiche 1857)
An mehreren Fundorten an der Küste wurden zahlreiche (insgesamt > 50 Ex.) Imagines von Hypolixus ornatus beobachtet, die sich bevorzugt an die Zweige von Tamarix (Tamariske) klammerten. Die Vermutung, dass die Art an Tamarix
lebt, wurde schnell verworfen, da die Tiere später auch an
krautigen Pflanzen gefunden wurden. Bei ersten Fraßtests wurde
Tamariske nicht, dafür jedoch verschiedene Chenopodiaceae
(Gänsefußgewächse) befressen. Wahrscheinlich entwickelt
sich die Art, wie viele verwandte Arten der Cleoninae an
Gänsefußgewächsen. Der Aufenthalt der Tiere auf Tamarix könnte mit der Überwinterung bzw. mit Wanderungsbewegungen in Zusammenhang stehen [W37.29]. Hypolixus ornatus wird in der Rüsselkäfer-Fauna von Zypern [Alziar 2007] abgebildet und trägt dort den Namen "Hypolixus pulvisculosus Boheman 1836". Nach Informationen von Lutz Behne (ZALF, Müncheberg) ist "pulvisculosus" ein jüngeres Synonym zu "ornatus". Wir gehen davon aus, dass es sich bei den Belegen von Zypern und denen von Rhodos um dieselbe Art handelt.
Larinus carinirostris Gyllenhal 1836
Die Art lebt im östlichen Mediterrangebiet an Carlina (Gold-/ Silberdistel, Asteraceae). Auf Rhodos wurde Larinus carinirostris
vorwiegend unter den Entwicklungspflanzen gefunden, da diese ihre
Blütenstände erst im Hochsommer entwickelt. Bei der
Präparation der mitgebrachten Belege stellte sich heraus, dass
einige der Imagines immatur waren. Dies stellt innerhalb der Larinus-Arten
eine Besonderheit dar, da sich die neue Generation in der Regel im
Hochsommer in den Köpfchen von Korbblütlern entwickelt und
diese noch im gleichen Jahr verlässt. Zur Klärung der
Beobachtung frisch geschlüpfter Imagines von Larinus cariniostris im April sind weitere Daten zur Lebensweise erforderlich [W37.30].
Lixus cribricollis Boheman 1836
Die Art wurde auf Rhodos sowohl an Fundort 14 (Theologos, Strand)
als auch an Fundort 19 (Apolakkia, Stausee) nachgewiesen. Während Lixus cribricollis an Fundort 19 als Imago auf Rumex
(Ampfer, Familie Knöterichgewächse - Polygonaceae) gefunden
wurde und sich offensichtlich zur Paarung und Eiablage auf den Pflanzen
aufhielt, wurden an Fundort 14 bei der Suche nach Apion-Arten an Emex spinosa
(L.) Campd. (Polygonaceae) Larven im Stängel absterbender Pflanzen
gefunden, aus denen nach einem Monat Tiere der neuen Generation von Lixus cribricollis schlüpften. Lixus cribricollis
ist also innerhalb der Knöterichgewächse oligophag. Bisher
waren nach Dieckmann [Dieckmann 1983] nur Arten der Gattung Rumex
als Entwicklungspflanze bekannt. Darüber hinaus zeigt der
zeitgleiche Fund der Art in unterschiedlichen Stadien der Entwicklung,
wie stark das Mikroklima verschiedener Biotope den Lebenszyklus und
damit auch die Nachweisbarkeit einer Art beeinflussen kann. Interessant
ist auch die Beobachtung, dass die Larven von Apion frumentarium (L.) (= A. miniatum auct.), welches an verschiedenen Fundorten (8, 14, 15) an Emex spinosa
gefunden wurde, sich in den Nodien (Knoten) der Sprossachsen
entwickeln, während die Internodien (Stängelabschnitte
zwischen den Knoten) gleichzeitig von Lixus cribricollis genutzt werden [W37.31] [W37.32].
Lixus junci Boheman 1836
Die Art konnte an Fundort 14 beobachtet werden. Die Imago hielt sich auf Beta vulgaris
L., der Wildform des Mangold, auf. Im Stängel der Pflanze, dem Ort
der Entwicklung der Larven, waren typische Eiablagespuren zu erkennen. Lixus junci
ist ein gutes Beispiel dafür, wie hilfreich botanische Kenntnisse
bei der Arterkennung im Gelände sind. Achtet man nur auf die
auffälligen Merkmale der Imago, so wäre aufgrund des
markanten weißen Seitenstreifens eine Verwechslung mit der
häufiger anzutreffenden Art Lixus albomarginatus möglich. Dieser lebt jedoch ausschließlich an Kreuzblütlern (Brassicaceae) und wurde auf Rhodos an FO 15 auf Cakile maritima Scop. (Europäischer Meersenf) gefunden [W37.33].
Orthochaetes (= Comasinus) sp.
Bei der Bodensuche und beim Abklopfen der Krautschicht wurden an verschiedenen Fundorten Imagines der Gattung Orthochaetes
gefunden. Besonders zahlreich (ca. 50 Exemplare) war die Art an Fundort
4 und 15, wo sie in Sandstrand- und Küstenfels-Biotopen gefunden
wurde. Bei Fraßtests mit Pflanzen aus der Nähe der
Fundstellen wurden Fabaceae (Schmetterlingsblütler) befressen.
Besonders gut wurden Lotus halophilus Boiss. & Spruner und Onobrychis sp. (Esparsette) angenommen. Ebenfalls an Fundort 15 wurde eine weitere Art der Gattung Orthochaetes gefunden, die möglicherweise an Arten der Asteraceae (Korbblütler) lebt [W37.22].
Pseudoprotapion elegantulum (Germar 1818)
Die Art ist nach Dieckmann [Dieckmann 1977] in größeren
Teilen Europas (fehlt in Nordeuropa), Vorder- und Mittelasien sowie
Sibirien und Nordafrika verbreitet. Die Entwicklung findet
ausschließlich an Arten der Gattung Onobrychis
(Esparsette) statt. Auf Rhodos wurden die Imagines an den noch nicht
blühenden Blütenständen einer mehrjährigen
Esparsetten-Art beobachtet. Die Weibchen fraßen Löcher in
die Blütenknospen und waren offensichtlich gerade bei der Eiablage
[W37.34].
Sitona syriacus Stierlin 1885
Bei der Bodensuche in einem Sandstrandbiotop (Fundort 15) [W37.35] mit artenreicher krautiger Sandküstenvegetation wurde Sitona syriacus gefunden. Bei der gezielten Suche nach den Flächen mit der größten Individuendichte konnte Sitona syriacus mit Lotus halophilus
Boiss. & Spruner als potentieller Entwicklungspflanze in
Zusammenhang gebracht werden. Die Imagines waren überwiegend
immatur (frisch geschlüpft). Einige Tiere gehörten zur
überwinterten Generation. Larven oder Puppen wurden nicht
beobachtet [W37.36].
Strophomorphus porcellus (Schoenherr 1832)
An Fundort 4 und 15 wurden bei der Bodensuche mehrere überwiegend immature Exemplare von Strophomorphus porcellus
gefunden. Die Tiere sind sehr lichtscheu und graben sich kurz nach
einer Störung in den Untergrund ein. Offensichtlich sind sie
nachtaktiv. An Fundort 7 wurden beim Graben im Boden eine Puppe und
zwei Larven gefunden. Die Puppe, deren Erdkokon zerstört worden
war, wurde bis zum Schlüpfen in angefeuchtetem Zellstoff
aufbewahrt. Ende April, circa drei Wochen nach dem Fund, schlüpfte
die Imago. Strophomorphus porcellus ist offensichtlich ein
Larvalüberwinterer, dessen Imagines von Mitte April bis in den
Herbst anzutreffen sind. Überwinterte Alt-Imagines wurden nicht
gefunden. Die "erdverbundene" Lebensweise und das Aussehen waren wohl
der Grund, warum der Erstautor die Art als "Schweinchen bzw. Ferkel"
(Bedeutung des lateinischen Wortes "porcellus") titulierte [W37.37].
Trichosirocalus urens (Gyllenhal 1837)
Die Arten der Gattung Trichosirocalus sind bis auf wenige, sogenannte "häufige" Arten (Trichosirocalus troglodytes, Trichosirocalus horridus) oft nur durch gezielte Suche am Boden zu finden. Auf Rhodos gelang der Nachweis von Trichosirocalus urens an mehreren Fundorten. Die Tiere wurden stets unter den bodennahen Blättern von Carlina graeca Boiss. (Asteraceae) gefunden. Larven oder Puppen wurden nicht gefunden. Trichosirocalus urens wird auch von weiteren Asteraceae gemeldet [Colonnelli 2004].
4. Diskussion
Die jahreszeitlichen Bedingungen
Für das Ziel der Erfassung
möglichst vieler Arten ist eine zweiwöchige Exkursion
grundsätzlich viel zu kurz, da sich der Zeitraum der optimalen
Nachweisbarkeit bezogen auf die verschiedenen Gruppen der
Rüsselkäfer über viele Monate verteilt. Auch stellte
sich heraus, dass viele Arten in der ersten Aprilhälfte gerade in
der "Larvalphase" sind, was einen Nachweis besonders dann erschwert,
wenn wenige oder keine Informationen zur Entwicklungspflanze bzw. zum
genutzten Pflanzenteil vorliegen. Findet man z. B. keine Imagines der
Gattung Otiorhynchus, obwohl das Vorkommen der Verwandtschaft
auf Rhodos bekannt ist, so müsste man zum Nachweis unkalkulierbar
große Mengen Erdreich "durchsuchen", um an Larven zu kommen, die
nur schwierig aufzuziehen sind. Aufgefundene Larven erfordern stets
einen hohen Zeitaufwand bei der Aufzucht, die spätere Zuordnung zu
einer Art ist jedoch nur auf diese Weise möglich. Für die
Erfassung der Hyperini war der Zeitraum hingegen nahezu optimal, da z.
B. bei vielen Donus-Arten nur in der Larvalphase Beobachtungen
zur Bindung an die Entwicklungspflanze möglich sind und die
Aufzucht der Larven relativ einfach ist [Dieckmann 1989]. Der Nachweis
von Exemplaren aus den artenreichen Taxa Apionidae und Ceutorhynchinae
hingegen war eher problematisch, da die neue Generation
überwiegend noch nicht geschlüpft war und daher viele Arten
– deren Vorkommen zu erwarten ist – nicht gefunden werden
konnten.
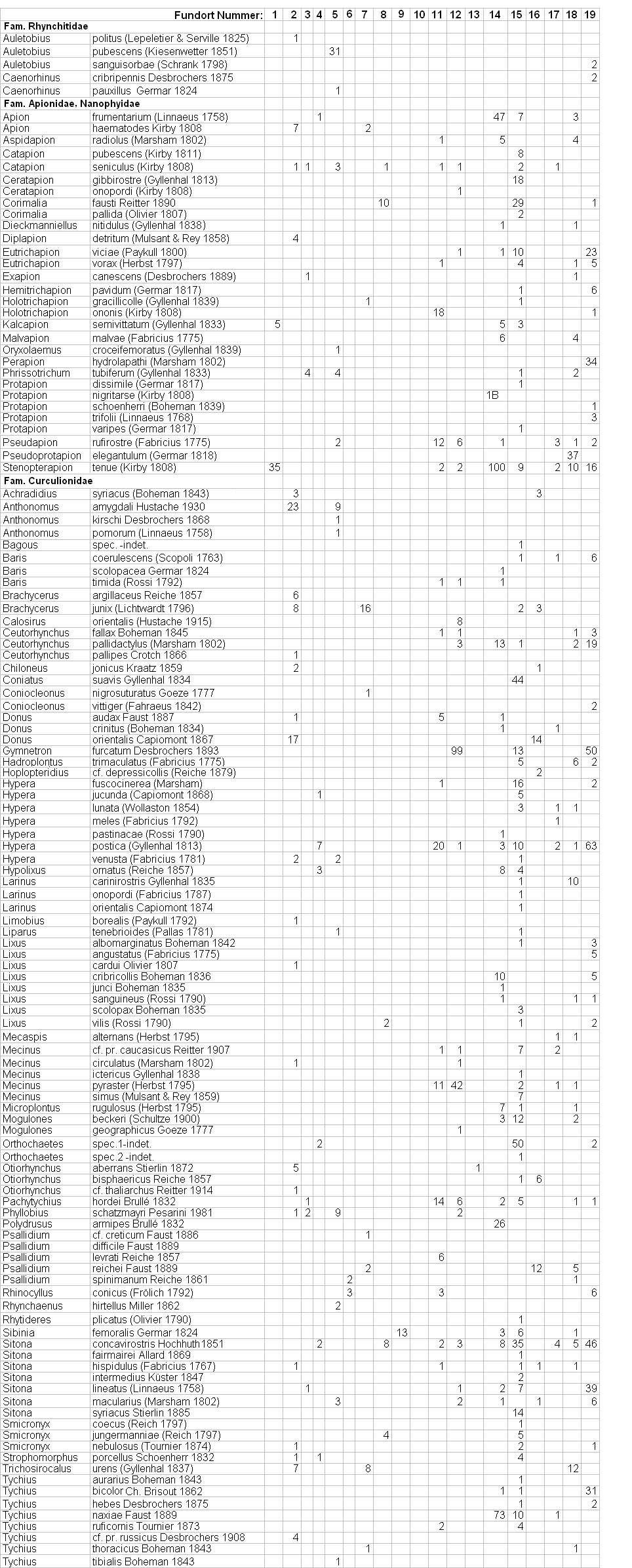
Liste der Arten von Rhodos
Insgesamt wird die
gemessen am Aufwand relativ geringe Zahl nachgewiesener Arten dadurch
"ausgeglichen", dass bewusst viel Zeit für die Fotodokumentation
und biologische Beobachtungen eingeplant wurde. Durch diese
Vorgehensweise kamen einige interessante Beobachtungen zur Lebensweise
sowie zur Biotop- und Entwicklungspflanzenbindung zustande. Über
die bloße Artenliste hinaus, stellen diese Bilddokumente und
Beobachtungen die eigentliche Grundlage unseres Beitrags dar.
Die auf Rhodos nachgewiesenen 127
Rüsselkäferarten können hier lediglich in einer
vorläufigen Artenliste zusammengefasst werden, da die
Determination einiger Taxa noch aussteht! [W37.Artenliste]
Determination der Arten
Da weder für Rhodos noch für
Griechenland oder die Türkei zusammenfassende Bestimmungsliteratur
vorliegt, haben wir auf die altbekannten und oft unzureichenden
Bestimmungswerke für den mittel- und westeuropäischen Raum
zurückgegriffen [Freude 1883] [Hoffmann 1950] [Hoffmann 1954]
[Hoffmann 1958] [Kippenberg 1981] [Tempère & Péricart
1989] oder verschiedene Gattungs-Monographien genutzt [Caldara 1990]
[Caldara 1998]. Darüber hinaus nutzten wir unsere privaten
Vergleichssammlungen und in schwierigeren Fällen die hervorragend
sortierte Sammlung des Deutschen Entomologischen Instituts
(Müncheberg). In diesem Zusammenhang muss erwähnt werden,
dass einige Taxa aus dieser bedeutenden Vergleichssammlung seit 15
Jahren ausgeliehen sind – offensichtlich also vom vorgeblichen
"Bearbeiter" nicht bearbeitet werden – und somit für andere
Wissenschaftler unzugänglich bleiben. Dies trifft z. B. auf die
Verwandtschaft um die Gattung Orthochaetes (= Comasinus)
zu. Die Belege dieser Taxa konnten wir daher nicht determinieren. Die
kritische Haltung entomologischer Museen gegenüber privaten
Sammlern sollte zuallererst dort zum Ausdruck gebracht werden, wo ihr
eigenes Material als "Dauerleihgabe" in privaten
Sammlungsschränken vermodert!
Danksagung
Unser Dank gilt den Kollegen Lutz Behne
(Eberswalde) und Roman Borovec (Nechanice), die uns mit Auskünften
über das Untersuchungsgebiet unterstützt haben. Herzlicher
Dank geht an Dr. Peter Stüben (Mönchengladbach) und Dr. Peter
Sprick (Hannover), die unsere Arbeit immer wieder hilfreich
unterstützt haben. Besonders bedanken wir uns bei Marion Brunner
(Berlin), die die uns auf der Exkursion begleitete und uns durch ihre
Mitarbeit viele interessante Beobachtungen ermöglichte. Vor Ort
hatten wir in Wolfgang Hock (Erfurt) fachkundigen Beistand bei der
Determination unserer Orchideenbilder.
5. Literatur
Alziar, Gabriel (19. August 2007): The Curculionoidea-Fauna of Cyprus. - Le Charançon: Catalogues & Keys, No. 3, ISSN 1864-0699, CURCULIO-Institute, Mönchengladbach.
Abbazzi P., Colonnelli E., Masutti L. & Osella G. (1994):
Coleoptera Polyphaga XVI (Curculionoidea). In: Minelli A., Ruffo S.
& La Posta S. (eds): Checklist delle specie della fauna italiana,
61. Calderini, Bologna, 1-68.
Caldara R. (1990):
Revisione tassonomica delle specie paleartiche del genere Tychius
Germar (Coleoptera Curculionidae). Memorie della Società
Italiana di Scienze Naturali e del Museo Civico di Storia Naturale di
Milano, 25: 51-218.
Caldara R. & O’Brien C. W. (1998): Systematics and
evolution of weevils of the genus Bagous. VI. Taxonomic treatment of
the species of the western Palearctic Region (Coleoptera
Curculionidae). Memorie della Società Entomologica Italiana, 76
(1997): 131-347.
Colonnelli E. (2004): Catalogue of Ceutorhynchini of the world, with a key to genera. Barcelona.
Dieckmann L. (1972): Beiträge zur Insektenfauna der DDR:
Coleoptera – Curculionidae: Ceutorhynchinae. Entomologische
Nachrichten und Berichte, 22, 3-128.
Dieckmann L. (1977): Beiträge zur Insektenfauna der DDR:
Coleoptera – Curculionidae (Apioninae). Entomologische
Nachrichten und Berichte, 27, 7-143.
Dieckmann L. (1983): Beiträge zur Insektenfauna der DDR:
Coleoptera – Curculionidae (Tanymecinae, Leptopiinae, Cleoninae,
Tanyrhynchinae, Cossoninae, Raymondionyminae, Bagoinae, Tanysphyrinae).
Entomologische Nachrichten und Berichte, 33, 257-381.
Dieckmann L. (1989): Die Zucht mitteleuropäischer Hyperini-Arten (Coleoptera, Curculionidae). Entomologische Nachrichten und Berichte, 33, 97-102.
Freude H., Harde K. W. & Lohse G.A. (1983): Die Käfer Mitteleuropas, 11, Goecke & Evers, Krefeld, 342.
Hoffmann A. (1950): Faune de France, 52,
Coléoptères Curculionides (Première Partie).
Librairie de la Faculté des Sciences, Paris, 1-486 pp.
Hoffmann A. (1954): Faune de France, 59,
Coléoptères Curculionides (Deuxième Partie).
Réimpression 1986, Fédération Francaise des
Sociétés de Sciences Naturelles, Paris, 487-1208 pp.
Hoffmann A. (1958): Faune de France, 62,
Coléoptères Curculionides (Troisième Partie).
Réimpression 1986, Fédération Francaise des
Sociétés de Sciences Naturelles, Paris, 1209-1841 pp.
Kippenberg H. (1981): 93. Familie: Curculionidae, pp. 102-310. In: Freude, H., Harde, K. W. & Lohse, G. A.: Die Käfer Mitteleuropas, 10, Goecke & Evers, Krefeld.
Tempère G. & J. Péricart (1989): Coleoptères
Curculionidae (Quatrième partie: Compléments). Faune de
France, 74: Fédération Francaise des
Sociétés de Sciences Naturelles, Paris, 1-534.
Adressen der Autoren
Christoph Bayer
Steilpfad 76
D-13509 Berlin / Germany
E-Mail: baris.bayer@arcor.de
Herbert Winkelmann
Attendorner Weg 39A
D-13507 Berlin / Germany
E-Mail: hyperiniwinkelmann@web.de
Friedhelm Bahr
Heinz-Luhnen-Straße 20
D-41751Viersen / Germany
E-Mail: Fried.Bahr@t-online.de
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}